СОРОСОВСКИЙ ОБРАЗОВАТЕЛЬНЫЙ ЖУРНАЛ №2 1997 Г.
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ВЗАИМОДЕЙСТВИЯ ИММУНОКОМПЕТЕНТНЫХ КЛЕТОК
В. Г. ГАЛАКТИОНОВ
Московский Государственный университет им. М.В. Ломоносова
ВВЕДЕНИЕ
Защитная реакция организма от патогена или любого другого чужеродного биологического материала осуществляется двумя системами иммунитета. В тех случаях, когда организм инфицируется бактериями, основная нагрузка падает на так называемую В-систему иммунитета. В состав этой системы входят костный мозг Ц основной источник клеток, продуцирующих специфические антитела и получивших название В-лимфоцитов (от англ. bone marrow - костный мозг), собственно В-лимфоциты и набор различных классов антител. Конечным результатом работы В-системы является накопление специфических антител, которые нейтрализуют бактерии или их токсины. Если же организм столкнулся с вирусной инфекцией, то в работу вступает Т-система иммунитета. Составляющими элементами Т-системы являются тимус (лимфоэпителиальный орган, расположенный за грудиной), различные субпопуляции тимусзависимых клеток (Т-лимфоцитов), антигенраспознающие рецепторы, находящиеся на поверхности этих клеток (Т-клеточные рецепторы - ТКР), а также группа регуляторных молекул (цитокинов). Одна из клеточных субпопуляций этой системы - Т-киллеры (цитотоксические Т-лимфоциты) являются основным компонентом антивирусного иммунитета. Т-киллеры контактируют с инфицированными вирусом клетками и разрушают их. Попавшие в кровоток вирусные частицы нейтрализуются антителами.
Изучая тонкие механизмы этих процессов, исследователи поняли, что ни В-клетки, ни Т-киллеры не в состоянии развить максимально эффективную реакцию самостоятельно. Для наиболее полного ее проявления необходима помощь со стороны других клеток. Эту вспомогательную функцию выполняют как фагоцитирующие клетки (макрофаги, дендритные клетки), так и Т-хелперы (от англ. helper - помощник). Именно через процесс взаимодействия различных типов иммунокомпетентных клеток формируется наиболее выраженный иммунный ответ.
Параллельно и вначале независимо от проблем взаимодействия клеток, ответственных за иммунный ответ, проводились исследования по генетической организации главного комплекса гистосовместимости (ГКГ). Этот комплекс генов был открыт в связи с пересадками (трансплантацией) тканей от одной особи вида другой. Отсюда и название комплекса.
До конца 60-х Ц начала 70-х годов было известно лишь одно свойство этого комплекса - контроль синтеза антигенов, вызывающих иммунную реакцию отторжения пересаженной ткани. Позднее при изучении генетического контроля силы иммунного ответа и особенно при анализе механизмов взаимодействия генетически отличающихся клеток был выявлен достаточно широкий спектр биологической активности комплекса. Под контролем ГКГ проходят такие иммунологические процессы, как регуляция силы гуморального (В-клеточного) и клеточного (Т-клеточного) иммунного ответа, обеспечение иммуногенности проникшего в организм антигена, селекция специфических Т-клеток в тимусе. И наконец, изучение участия генов ГКГ во взаимодействии иммунокомпетентных клеток привело к пониманию одного из существенных явлений иммунитета - распознавания антигена Т- и В- клетками.
ГЛАВНЫЙ КОМПЛЕКС ГИСТОСОВМЕСТИМОСТИ
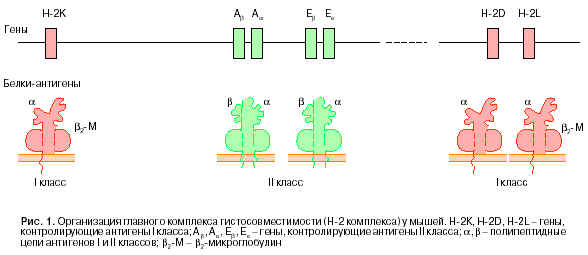
ГКГ у мышей представляет собой группу близкосцепленных генов, локализованных в 17-й хромосоме. У человека этот комплекс располагается в 6-й хромосоме. Гены Н-2К, Н-2D и Н-2L ответственны за синтез антигенов I класса. Гены Аb, Аa, Eb, Ea обеспечивают образование полипептидных цепей, которые формируют антигены II класса. Известны две формы таких антигенов АbАa и EbEa (рис. 1).
Антигены I класса являются гликопротеинами с молекулярной массой 44000. С основной, тяжелой a--цепью этого антигена нековалентно связан b2-микроглобулин (b2-М), имеющий близкую гомологию с иммуноглобулинами. Антигены II класса построены из двух полипептидов (b- и a- цепей). Антигены I и II класса, а также b2-микроглобулин по характеру строения и высокой гомологии последовательности аминокислотных остатков с иммуноглобулинами входят в состав единого суперсемейства. Этот факт сам по себе примечателен. Он указывает на то, что роль ГКГ состоит не только в маркировке индивидуальности, но и в реакциях иммунологической природы.
ВЗАИМОДЕЙСТВИЕ ГЕНЕТИЧЕСКИ ИДЕНТИЧНЫХ (СИНГЕННЫХ) КЛЕТОК
Проблема межклеточных взаимодействий возникла в биологии достаточно давно. Исследователям, занимающимся изучением вопросов клеточной биологии, хорошо известно взаимное влияние различных типов клеток в таких процессах, как дифференцировка клеток, эмбриональное развитие, регуляция роста тканей. Однако в иммунологии изучения вопросов клеточного взаимодействия как одного из еханизмов осуществления иммунологических функций до середины 60-х годов не проводилось.
Казалось, что основные клетки Т- и В-систем работают автономно. Первые из них - для реализации клеточного типа ответа, вторые Ц гуморального. В 1963 году появилась публикация Claman, в которой было показано, что для полноценной продукции антител необходима кооперация по крайне мере двух типов клеток: Т- и В-лимфоцитов. Исследователи провели простые, но при этом крайне демонстративные опыты. Летально облученным мышам, лишенным собственных иммунокомпетентных клеток, вводили либо только клетки тимуса (источника Т-лимфоцитов), либо клетки костного мозга (источника В-клеток), либо смесь этих клеток. В первых двух случаях регистрировали лишь следы антител после введения антигена. В третьем варианте опытов величина ответа была значительно выше суммы ответов двух предыдущих групп. Налицо был явный синергический эффект. Именно синергизм при кооперации двух типов клеток вызвал у иммунологов особый интерес. После работы Claman иммунологические журналы были переполнены статьями, касающимися различных сторон клеточной кооперации. При всей впечатляющей силе первой публикации в работе был существенный недостаток. Опыты проводили на животных (in vivo), и в силу этого не было возможности учесть всех участников клеточного взаимодействия.
Несколько позднее Mosier и Coppleson (1968) провели эксперименты с культурой клеток. Получив очищенные от посторонних клеток популяции Т-, В-лимфоцитов и макрофагов (МФ), они стали комбинировать между собой различные типы клеток in vitro. Из всех возможных вариантов: Т-лимфоциты плюс В-лимфоциты, Т-лимфоциты плюс макрофаги, В-лимфоциты плюс макрофаги и, наконец, Т-лимфоциты плюс В-лимфоциты плюс макрофаги Ц продукция антител наблюдалась только при сочетании трех типов клеток. Стало очевидно, что полноценное развитие гуморального иммунного ответа требует не двух, а по крайней мере трех типов клеток. Функция каждого клеточного типа в антителопродукции строго предопределена. Макрофаги и другие фагоцитирующие клетки поглощают, перерабатывают и экспрессируют антиген в иммуногенной, доступной для Т- и В- лимфоцитов форме. Т-клетки, а точнее, одна из субпопуляций этих клеток, так называемых Т-хелперов (помощников), после распознавания антигена начинает продукцию цитокинов, обеспечивающих помощь В-клеткам. Эти последние клетки, получив специфический стимул от антигена и неспецифический от Т-клеток, вступают в основной эффекторный процесс - продукцию антител (рис. 2).
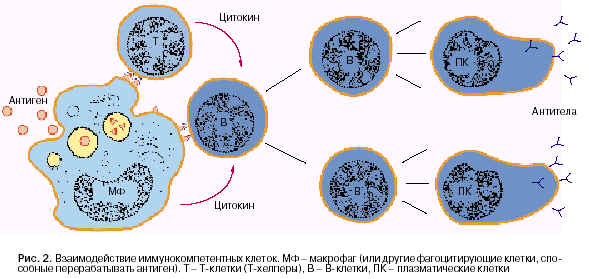
При формировании клеточного типа иммунного ответа также необходима кооперация различных типов клеток. Предшественник клеточного ответа, например Т-киллер, после распознавания антигена испытывает на себе действие Т-хелперов и вспомогательных клеток (фагоцитирующих мононуклеаров). Только после взаимодействия предшественника с этими клетками появляется возможность его развития до зрелой эффекторной клетки. Внимательное изучение механизмов кооперации иммунокомпетентных клеток привело к формированию одного из наиболее перспективных направлений исследования, связанного с изучением роли гуморальных факторов в реализации клеточного взаимодействия. Современное название этих факторов - цитокины. Сегодня известны структура и функция около 20 таких цитокинов. Более того, для многих из них получены генно-инженерные аналоги, повторяющие активность природных соединений. В табл. 1 в качестве примера представлены некоторые из известных цитокинов, осуществляющих реальную связь между клетками - продуцентами и мишенями, что и вскрывает один из аспектов клеточного взаимодействия.

ВЗАИМОДЕЙСТВИЕ ГЕНЕТИЧЕСКИ ОТЛИЧАЮЩИХСЯ КЛЕТОК
C середины 60-х годов начало формироваться направление, задача которого - выяснение генетических механизмов, регулирующих силу иммунного ответа. Были обнаружены гены иммунного реагирования (Ir-гены - immune response genes), которые находятся в ГКГ. Сразу после локализации генов в хромосоме Ir возник вопрос о типе клеток, в которых проявляется активность этих генов. Необходимость решения данного вопроса привела к созданию комбинированных клеточных культуральных систем, составленных от животных с разной способностью к иммунному ответу на те или иные антигены. Как часто бывает в науке, используемая система взаимодействия генетически отличающихся (несингенных или аллогенных) клеток оказалась полезной не только для ответа на выдвинутый вопрос. Она принесла пользу в решении проблемы антигенного распознавания Т-лимфоцитами. Дело в том, что до середины 80-х годов природа антигенраспознающих рецепторов Т-клеток (ТКР Ц Т-клеточный рецептор) оставалась неизвестной. И только совместные усилия клеточных и молекулярных иммунологов привели к пониманию характера антигенного распознавания тимусзависимыми клетками. Первые опыты, заставившие думать об особом характере ТКР, отличном от антигенраспознающих рецепторов В-клеток (поверхностных иммуноглобулинов - sIg), были проведены с культурой клеток. Так, например, в опытах in vitro была предпринята попытка оценить реакцию Т-клеток (конкретно Т-хелперов) на чужеродный антиген, который был ассоциирован с макрофагами. Выяснилось, что при генетической идентичности между макрофагом и Т-хелперами последние развивают сильную ответную реакцию. Ситуация менялась, если Т-клетки генетически отличались от макрофагов. Во всех случаях, когда различия касались генов, контролирующих антигены II класса, Т-хелперы оставались пассивными. Подобная инертность показалась странной. Действительно, если чувствительные к определенному антигену Т-хелперы распознают только тот антиген, к которому они были сенсибилизированы, им все равно, с каким макрофагом взаимодействовать. Однако факты говорили об обратном. Оставалось предположить, что Т-хелперы распознают не только чужеродный антиген на поверхности макрофагов, но и свои собственные антигены II класса, представленные на тех же макрофагах (рис. 3).
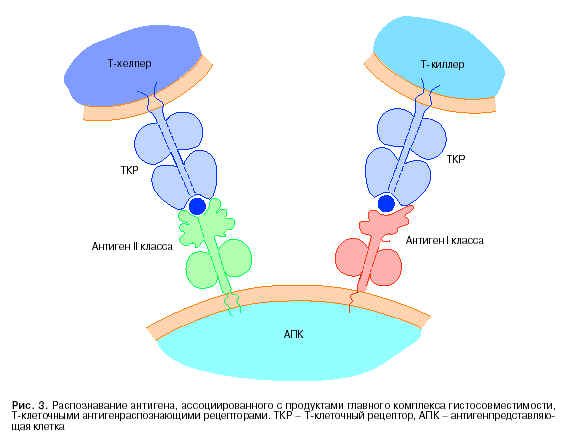
Если Т-хелперы распознают комплекс чужеродного, экзогенного антигена с антигенами II класса ГКГ, то генерация цитотоксических Т-лимфоцитов (Т-киллеров) связана с ограничениями по антигенам I класса. К такому выводу пришли Zinkernagel и Doherty после проведения серии исследований по индукции Т-киллеров под контролем со стороны ГКГ. Специфически сенсибилизированные к определенному вирусу Т-киллеры способны разрушать в условиях in vitro генетически идентичные (сингенные) клетки-мишени, зараженные тем же вирусом. Смена одного вируса на другой отменяла реакцию лизиса, что указывало на специфичность анализируемых Т-киллеров. Однако эта специфичность была ограничена антигенами I класса ГКГ (Н-2К или Н-2D). Реакция на специфический вирус развивалась только при идентичности по антигенам I класса ГКГ между премированными Т-клетками и клетками-мишенями, зараженными соответствующим вирусом (см. рис. 3).
Итак, Т-киллеры, так же как и Т-хелперы, распознают не собственно чужеродный антиген, но его комплекс с продуктами ГКГ: Т-помощники Ц комплекс с антигенами II класса, Т-киллеры Ц с антигенами I класса. В схеме генерация и последующее функциональное проявление двух типов клеток выглядят следующим образом. Проникший в организм чужеродный антиген (например, вирус) захватывается и перерабатывается фагоцитирующей клеткой. В результате внутриклеточного процессинга фрагменты антигена выходят на клеточную поверхность, где и образуют комплекс с антигенами I или II класса. Включение в комплекс антигенов ГКГ и является тем генетически обусловленным ограничением, которое не позволяет развиваться ответу в тех случаях, когда в комплекс включены антигены иного генотипа.
Клеточная феноменология распознавания комплексного антигена Т-хелперами и Т-киллерами указывает лишь на двойной характер такого распознавания (распознавания ''своего'' и ''чужого''), но ничего не говорит о природе собственно антигенраспознающих структур Т-клеток. История изучения этих структур полна ошибок, спекуляций и различного рода общих суждений. Действительно, явление взаимодействия иммунокомпетентных клеток в условиях ответа на антиген ясно указывало на особые свойства рецепторов Т-клеток, отличных от аналогичных поверхностных структур В-клеток, которым не требуются антигены ГКГ для реализации процесса распознавания. Отсутствие же конкретных данных по молекулярной организации антигенных рецепторов Т-клеток и приводило к умозрительным построениям, многие из которых так и не нашли экспериментального подтверждения. Только в середине 80-х годов была наконец понята молекулярная природа Т-клеточных рецепторов. Они представляют собой гликопротеины клеточной поверхности, построенные из двух полипептидных цепей: a- и b- . Каждая цепь включает два домена: вариабельный (V) и константный (С). Антигенраспознающий центр, так же как и у антител, формируется при взаимодействии V-доменов каждой из цепей. Сходство с антителами касается и организации генов, контролирующих синтез рецепторов Т- клеток. Между двумя типами рецепторов имеется значимая гомология по последовательности аминокислотных остатков в полипептидах. Предполагается, что эволюционно Т-клеточные рецепторы явились предшественниками антител. Черты сходства между этими структурами позволили объединить их наряду с другими близкими белками в единое суперсемейство.
ТИМУС - ШКОЛА ОБУЧЕНИЯ НЕЗРЕЛЫХ Т-КЛЕТОК
В середине 60-х годов была установлена ключевая роль тимуса в формировании пула зрелых Т-клеток. Позднее стало очевидно, что именно в тимусе происходят процессы дифференцировки Т-клеток на субпопуляции (Т-киллеры, Т-хелперы, Т-супрессоры). И наконец, в середине 80-х годов иммунологи поняли, каким образом в тимусе Т-клетки приобретают свое главное свойство - способность к двойному распознаванию. Путь созревания и ''обучения'' предшественников Т-клеток в тимусе выглядит следующим образом (рис. 4). Незрелые клетки (пре-Т-лимфоциты), лишенные как маркеров дифференцировки, так и Т-клеточных, антигенраспознающих рецепторов (ТКР), мигрируют по предначертанному пути из костного мозга в субкапсулярную зону органа. Здесь осуществляются первые шаги внутритимусной дифференцировки лимфоцитов. В результате на поверхности клеток начинается одновременная экспрессия двух маркеров дифференцировки (CD4 Ц маркер Т-хелперов и CD8 - маркер Т-киллеров). Однако у тимоцитов субкапсулярной зоны еще отсутствуют ТКР. Их умеренная экспрессия начинается позднее, после миграции клеток в кору. С момента появления антигенных рецепторов начинается основной этап обучения. Из множества пришедших в кору клеток успех в дальнейшем развитии сопутствует только тем, чьи рецепторы способны взаимодействовать с антигенами ГКГ, обильно представленными на стромальных клеточных элементах. Те клетки, рецепторы которых в силу специфической конформации активного центра неспособны взаимодействовать с антигенами ГКГ, погибают.
Селекция различных клонов лимфоцитов - основное событие, разворачивающееся в одном из центральных органов иммунитета. Отбор клеток по способности распознавать свои собственные антигены является определяющим условием дальнейшего внутритимусного развития Т-лимфоцитов. В тех случаях, когда распознаются антигены I класса, лимфоциты выбирают путь развития в сторону Т-киллеров (маркер CD8). Если же распознавание связано с антигенами II класса, провзаимодействовавшие тимоциты трансформируются в Т-хелперы (маркер CD4).
Предполагается, что узнавание ''своего''осуществляется не всем антигенраспознающим центром ТКР, а лишь его частью. Оставшаяся часть центра будет взаимодействовать с возможными в будущем чужеродными антигенами. Именно тогда принцип двойного распознавания ''своего'' и ''чужого'' найдет свое реальное проявление. Хотелось бы сказать несколько слов об иммунологической загадке прошлого. Долгое время оставалось непонятным, почему в тимусе гибнут более 90% поступающих из костного мозга ранних предшественников Т-клеток. Известный австралийский иммунолог Ф. Бернет предполагал, что в тимусе происходит гибель тех лимфоцитов, которые способны к аутоиммунной агрессии. Иначе говоря, тимус выступал в качестве заслона нежелательным или даже ''вредным'' клонам. Сегодня в данной проблеме имеется определенная ясность. Основная причина столь массовой гибели связана с жесткостью селекционных процессов - положительным отбором только тех клеток, которые способны реагировать со своими собственными антигенами ГКГ. Все остальные не прошедшие контроля на специфичность погибают.
ЗАКЛЮЧЕНИЕ
Изучение генетического контроля взаимодействия иммунокомпетентных клеток привело к пониманию наиболее скрытых механизмов работы имунной системы, и в первую очередь к выяснению природы антигенраспознающих рецепторов Т-клеток. Весь путь познания в данном случае можно разделить на несколько этапов.
1. Определение тимуса как центрального органа иммунитета, где закладываются основы клеточного типа реагирования.
2. Формирование представлений о двух системах специфической защиты: Т- и В-системах иммунитета.
3. Установление взаимодействия различных типов клеток при развитии иммунного ответа.
4. Открытие в системе взаимодействия генетически отличающихся клеток способности Т-клеточных рецепторов к двойному распознаванию.
5. Выяснение молекулярной природы Т-клеточного антигенраспознающего рецептора.
6. Установление селекционирующей роли тимуса Ц места формирования специфических Т-клеток, способных к двойному распознаванию: собственных антигенов гистосовместимости и комплексированных с ними чужеродных антигенов.
ЛИТЕРАТУРА
1. Ройт Р. Основы иммунологии. М.: Мир, 1991.
2. Иммунология / Под ред. У. Пола. М.: Мир, 1988. Т. 1Ц3.
3. Галактионов В.Г. Графические модели в иммунологии. М.: Медицина, 1986.
Вадим Геллиевич Галактионов, доктор биологических наук, профессор, сотрудник Института биологии развития им. Н.К. Кольцова РАН. Область научных интересов: генетика и эволюция иммунитета. Автор более 120 работ и трех монографий.