СОРОСОВСКИЙ ОБРАЗОВАТЕЛЬНЫЙ ЖУРНАЛ №5 1997
ЗАГАДКИ ТИМУСА. ВОЗРАСТ И ИММУНИТЕТ.
И.С. ФРЕЙДЛИН
САНКТ-ПЕТЕРБУГРСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ ИНСТИТУТ ИМ. ПАВЛОВА
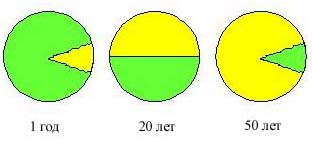
В центральной части грудной клетки, непосредственно за грудиной, расположен один из самых таинственных органов иммунной системы – тимус или вилочковая железа. У новорожденного младенца тимус уже полностью развит и в течение первого года жизни достигает максимальных размеров 25 см3. Вес тимуса шестимесячного ребенка достигает 20 г, что составляет 0,5% веса тела. Его сложная структура и напряженное функционирование сохраняются в первые 2–3 года жизни. К 20 годам половина функционирующей ткани тимуса замещается жировой тканью. К 50–60 годам инволюция тимуса завершается (рис. 1). Этот орган представляет собой пример чрезвычайно раннего старения и отмирания. В связи с этим возникают вопросы: какие функции выполняет тимус в детском организме и нужен ли тимус во взрослом организме или организм может обойтись без тимуса?
Рис. 1. Возрастная инволюция тимуса. При сохранении размеров органа объем истинно тимической ткани (зеленый цвет) прогрессивно снижается в первые годы жизни и почти полностью замещается жировой тканью (желтый цвет) к 50 годам
ТИМУС - МЕСТО СОЗРЕВАНИЯ Т-ЛИМФОЦИТОВ
Роль тимуса была убедительно показана при изучении заболевания, получившего название “синдром ДиДжорджи (DiGeorge)”, при котором генетически детерминированное недоразвитие этого органа приводит к отсутствию одной из популяций лимфоцитов – Т-лимфоцитов. У детей с таким врожденным иммунодефицитом проявлялась повышенная чувствительность к вирусным, грибковым и некоторым бактериальным инфекциям [1].
Т-лимфоциты называются так, поскольку их накопление и созревание происходит в тимусе. Незрелые предшественники Т-лимфоцитов из костного мозга мигрируют в тимус, где получают название тимоциты” и находят наиболее благоприятное микроокружение для дальнейшего развития. В наружном слое тимуса, который называют корковым, строму (основу) ткани составляют эпителиальные клетки, имеющие множество отростков. Своими отростками эпителиальные клетки как бы окружают, обнимают незрелые тимоциты, за что и получили образное название “клетки-няньки”. По мере созревания тимоциты продвигаются из коркового слоя тимуса в глубину и попадают в “мозговой” слой, где встречаются с другими клетками стромы тимуса: макрофагами, дендритными клетками, фибробластами. Все эти клетки вступают в контакты с мигрирующими тимоцитами и влияют на них своими поверхностно расположенными молекулами – антигенами и своими продуктами: гормонами и цитокинами. На поверхностных мембранах всех перечисленных клеток экспрессированы антигены тканевой совместимости, с которыми комплексируются различные антигенные пептиды, в частности это могут быть фрагменты аутоантигенов [2]. Эпителиальные клетки тимуса продуцируют и секретируют белки и пептиды, получившие собирательное название “тимусные гормоны”. Среди тимусных гормонов отдельные молекулы контролируют пролиферацию, дифференцировку и функции тимоцитов. Они могут также усиливать функции зрелых Т-лимфоцитов, что позволяет использовать препараты очищенных экстрактов тимуса в клинике для лечения иммунодефицитов с выраженными дефектами Т-клеток.
Контактируя с эпителиальными и другими клетками стромы тимуса, каждый тимоцит последовательно получает сигналы: пролиферации, изменения поверхностного фенотипа, перестройки (реаранжировки) генов для приобретения Т-лимфоцитами широкого “репертуара” специфичностей рецепторов, позитивной и негативной селекции, функционального созревания. Эти процессы необходимы для превращения незрелого тимоцита в функционально полноценный Т-лимфоцит. Все процессы происходят на территории тимуса, только в различных его слоях и при участии разных стромальных клеток.
В тимусе идет постоянная пролиферация тимоцитов: из общего количества 2*10 8 тимоцитов 20– 25% (5*10 7) клеток заново образуются ежедневно при их делении. Но самое удивительное, что только 2–5%, то есть 1*10 6 из них, в виде зрелых Т-лимфоцитов ежедневно покидают тимус, поступая в кровь и расселяясь в лимфоидных органах. Это значит, что 50*10 6 (95–98%) тимоцитов ежедневно погибают в тимусе, а выживают в многоэтапном процессе развития и селекции лишь 2–5% клеток (рис. 2).
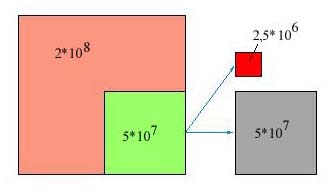
Рис. 2. В тимусе идет пролиферация тимоцитов, ежедневный прирост количества тимоцитов составляет 20–25% (зеленый цвет). Из них только 2–5% ежедневно покидают тимус в виде зрелых Т-лимфоцитов (красный цвет), а 95–98% погибают в тимусе (серый цвет)
СЕЛЕКЦИЯ НУЖНЫХ ТИМОЦИТОВ В ТИМУСЕ
Основная функция зрелых Т-лимфоцитов – это распознавание чужеродных антигенных пептидов в комплексе с собственными антигенами тканевой совместимости на поверхности вспомогательных (антигенпрезентирующих) клеток или поверхности любых клеток-мишеней организма [2]. Для выполнения этой функции Т-лимфоциты должны быть способны распознавать собственные антигены тканевой совместимости, специфичные для каждого индивидуума. Одновременно Т-лимфоциты не должны распознавать аутоантигенные пептиды самого организма, связанные с собственными антигенами тканевой совместимости. Между тем в процессе перестройки (реаранжировки) генов созревающих тимоцитов некоторые из них приобретают рецепторы Т-клеток (РТК), специфичные именно в отношении антигенных пептидов самого организма, то есть аутоантигенных пептидов. В связи с этим в тимусе одновременно с процессами пролиферации и созревания тимоцитов идут процессы их селекции – отбора нужных Т-лимфоцитов (рис. 3).
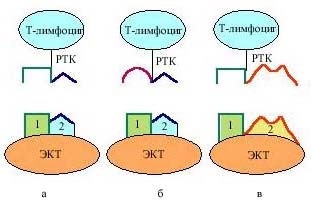
Рис. 3. Позитивная и негативная селекция Т-лимфоцитов в тимусе: а – Т-лимфоцит, способный с помощью своего рецептора распознать собственные антигены тканевой совместимости (1), экспрессированные на эпителиальных клетках тимуса (ЭКТ), проходит позитивную селекцию и вступает в дальнейшее развитие; б – Т-лимфоцит, неспособный распознать собственные антигены тканевой совместимости (1) на поверхности ЭКТ, не проходит позитивную селекцию и получает сигнал запрограммированной смерти – апоптоза; в – Т-лимфоцит, способный распознать пептидные фрагменты аутоантигенов (3) на поверхности антигенпрезентирующих клеток (АПК) в комплексе с собственными антигенами тканевой совместимости (1), на этапе негативной селекции получает сигнал запрограммированной смерти – апоптоза. РТК – рецептор Т-клеток, 2 – пептидные фрагменты чужеродных антигенов в комплексе с 1.
Селекция тимоцитов проходит в два этапа. После того как на индивидуальном тимоците экспрессируется РТК его уникальной специфичности, клетка вступает в этап позитивной селекции. Для того чтобы выжить и вступить в следующие этапы развития, тимоцит должен проявить способность распознавать собственные антигены тканевой совместимости, экспрессированные на эпителиальных клетках коры тимуса. Существуют сотни разных вариантов антигенов тканевой совместимости, из которых лишь малая часть экспрессирована на клетках данного индивида. Из широчайшего “репертуара” специфичностей РТК лишь немногие подойдут для распознавания индивидуального набора антигенов тканевой совместимости данного организма. Тимоциты с такими подходящими РТК получают сигнал дальнейшей дифференцировки. Они отобраны на этапе позитивной селекции и вступают в следующий этап (рис. 3, а ).
На границе коркового и мозгового слоев тимуса созревающие тимоциты встречаются с дендритными клетками и макрофагами. Профессия этих клеток – презентация антигенных пептидов в комплексе с собственными антигенами тканевой совместимости для распознавания Т-лимфоцитами. В данном случае эти клетки представляют пептиды самого организма – фрагменты аутоантигенов, которые могут заноситься в тимус с током крови. В отличие от зрелого Т-лимфоцита, который при встрече с антигенным пептидом, специфичным для его РТК, получает сигнал активации, незрелые тимоциты в тимусе при распознавании специфичных для их РТК антигенных пептидов получают сигнал генетически запрограммированной смерти – апоптоза [3]. Таким образом идет негативная селекция аутореактивных Т-лимфоцитов, которые подвергаются делеции (рис. 3, в ). В результате позитивной и негативной селекции из тимуса в кровоток и лимфоидные органы поступают только такие Т-лимфоциты, которые несут РТК, способные распознавать собственные молекулы тканевой совместимости в комплексе с пептидными фрагментами чужеродных белков и неспособные распознавать их в комплексе с аутоантигенными пептидами. Такие зрелые Т-лимфоциты берут на себя основные защитные функции в противовирусном и противоопухолевом иммунитете. Кроме того, они выполняют важные регуляторные функции, контролируя силу и форму любого специфического иммунного ответа [4]. Итак, необходимость тимуса в детском организме не вызывает сомнений.
ВОЗРАСТНАЯ ИНВОЛЮЦИЯ ТИМУСА
Вместе с тем не вызывают сомнения объективные данные, свидетельствующие об очень быстрой возрастной инволюции тимуса, то есть об утрате тимуса с возрастом. В течение первых 50 лет жизни ежегодно теряется по 3% истинно тимической ткани, которая постепенно замещается жировой и соединительной тканью. Соответственно снижается и продукция Т-лимфоцитов. Самая высокая продукция Т-лимфоцитов сохраняется до двух лет, а затем быстро падает. Возрастная инволюция тимуса происходит не только у человека. У мыши, например, к 24-месячному возрасту продукция Т-клеток составляет 0,7% уровня их продукции у новорожденной мыши. К этому возрасту у мыши происходит почти полная редукция тимуса: теряется и структура органа, и его функция. Однако следует отметить, что количество Т-лимфоцитов в циркуляции сохраняется на достигнутом уровне. Дело в том, что значительную часть популяции Т-лимфоцитов составляют долгоживущие клетки, которые не нуждаются в постоянном обновлении. Поэтому численность Т-клеток может поддерживаться во взрослом организме и при отсутствии тимуса. Более того, зрелые Т-лимфоциты подвергаются так называемой клональной экспансии, то есть избирательной пролиферации в ответ на встречу со своим антигеном, за счет чего их численность возрастает. После создания пула периферических Т-лимфоцитов утрата тимуса уже не приводит к катастрофическому снижению иммунитета. В пользу этого говорят результаты иммунологического обследования взрослых людей и мышей, перенесших тимэктомию [5].
Из всех органов иммунной системы только для тимуса характерна возрастная инволюция. Костный мозг не претерпевает подобных возрастных изменений, если не считать накопления жировых отложений. Не наблюдается возрастной инволюции ни селезенки, ни лимфоузлов. Дифференцировка гранулоцитов и моноцитов с возрастом даже усиливается. Увеличивается с возрастом количество естественных киллеров – больших гранулярных лимфоцитов, дифференцировка которых не связана с тимусом. Можно заключить, что в организме сохраняется воспроизводство всех остальных иммунокомпетентных клеток, которые не являются долгоживущими, выполняют функции эффекторов и тратятся постоянно в борьбе с болезнетворными микробами.
В отличие от этого необходимость в генерации новых Т-лимфоцитов снижается с возрастом. Первичные контакты с инфекционными агентами происходят в основном в первые годы жизни, когда и формируются Т-клетки памяти. Т-лимфоциты памяти у людей живут более 20 лет. В дальнейшем поступление новых патогенов становится более редким событием. Содержание организмом целого тимуса с его энергетической емкостью становится нецелесообразным, и тимус подвергается инволюции, так как его поддержание, репарация дорого обходятся организму, в то время как этот орган становится ненужным [5]. Однако отсутствие тимуса у взрослых людей может быть причиной того, что заражение их “детскими” вирусами – возбудителями кори или ветрянки приводит к развитию значительно более тяжелых, чреватых осложнениями заболеваний по сравнению с корью или ветрянкой у детей. Не зря именно в течение первых трех лет жизни, то есть в период расцвета функций тимуса, врачи стараются вакцинировать ребенка против всех наиболее опасных и частых инфекционных заболеваний: туберкулеза, полиомиелита, дифтерии, столбняка, коклюша, кори. В этом возрасте иммунная система организма, как правило, отвечает на введение вакцины (убитых или ослабленных возбудителей, их антигенов, обезвреженных токсинов) выработкой активного иммунитета – формированием клонов клеток памяти, в том числе долгоживущих Т-клеток памяти. При наличии такого клона организму не страшна встреча с болезнетворным микробом: РТК тут же распознают “запомнившиеся” антигены, получат сигнал клональной экспансии (пролиферации), активации и начнут выполнять свои защитные функции, что ведет к элиминации возбудителя и нейтрализации его токсинов.
При отсутствии тимуса его функции могут частично выполнять какие-то участки лимфоидных тканей, где идет созревание Т-лимфоцитов. Наиболее ярким примером компенсации функций отсутствующих Т-лимфоцитов могут служить так называемые голые (nude) мыши. У таких мышей имеется сочетание двух генетических дефектов: дефекта эпителия кожи, ведущего к отсутствию волосяного покрова, и недоразвития тимуса, ведущего к отсутствию Т-лимфоцитов. У таких мышей компенсаторно повышено количество естественных киллеров, которые способны продуцировать и секретировать один из важнейших защитных цитокинов – гамма-интерферон. При наличии в организме Т-лимфоцитов они являются основными продуцентами гамма-интерферона. При их отсутствии эту важную защитную функцию берут на себя другие клетки – естественные киллеры, развитие которых протекает без участия тимуса [1].
ЗАКЛЮЧЕНИЕ
Подводя итоги, следует напомнить, что тимус наряду с костным мозгом относится к центральным органам иммунной системы. Костный мозг дает начало всем росткам кроветворения: из единой стволовой полипотентной клетки костного мозга происходят и эритроциты, и тромбоциты, и гранулоциты, и моноциты, и лимфоциты. В отличие от костного мозга тимус узко специализирован на продукции Т-лимфоцитов. Особенность процессов созревания Т-лимфоцитов по сравнению с другими лейкоцитами заключается в необходимости отбора среди них клеток, способных распознавать собственные антигены тканевой совместимости. Из числа отобранных по этому признаку Т-лимфоцитов нужна дополнительная отбраковка клеток, способных распознать аутоантигенные пептиды в комплексе с собственными антигенами тканевой совместимости. На этих процессах селекции тимоцитов специализируются стромальные клетки тимуса: эпителиальные клетки, дендритные клетки, макрофаги. В работе тимуса много лишних затрат: активная пролиферация тимоцитов сопровождается 95%-ной их гибелью при прохождении двух ступеней селекции. Созданный с большими энергетическими затратами пул зрелых периферических Т-лимфоцитов включает относительно долгоживущие клетки, способные отвечать клональной экспансией (пролиферацией) на встречу с антигеном. Поэтому возрастная инволюция тимуса не приводит к катастрофическому снижению иммунитета. Кроме того, иммунная система располагает некоторыми компенсаторными возможностями замещения отдельных функций недостающих Т-лимфоцитов.
ЛИТЕРАТУРА
1. Ройт А . Основы иммунологии. М.: Мир, 1991.
2. Абелев Г.И . Основы иммунитета // Соросовский Образовательный Журнал. 1996. № 5. С. 4–10.
3. Агол В.И . Генетически запрограммированная смерть клеток // Там же. 1996. № 6. С. 20–24.
4. Janeway Ch ., Travers P . Immunobiology. L.: Curr. Biol. Ltd., 1994.
5. George A. , Ritter M . Thymic Involution with Ageing: Ob- solescence or Good Housekeeping? // Imm. Today. 1996. Vol. 17, № 6. P. 267–272.
Ирина Соломоновна Фрейдлин, доктор медицинских наук, профессор кафедры микробиологии, вирусологии и иммунологии Санкт-Петербургского государственного медицинского университета им. И.П. Павлова, руководитель отдела иммунологии НИИ экспериментальной медицины РАМН. Область научных интересов – фундаментальная и прикладная иммунология. Автор более 200 научных работ, в том числе пяти монографий, соавтор двух учебников.